Factors Affecting Cold Hardiness Development
FGV-00143 View this publication in PDF form to print or download.
Winter survival of perennial plants is a common problem for amenity, horticultural and agronomic crops in Alaska since winters here can be both severe and long. Native plants have developed mechanisms to deal with winters in Alaska, but efforts to import plants from more southern locations often prove disappointing. This report summarizes recent research on this subject. The majority of the work was conducted by Dr. L. J. Klebesadel in Palmer. The discussion includes a brief outline of environmental conditions and associated stresses on plants, the importance of latitude of origin, plant mechanisms for surviving winter stresses, and the potential for management to influence survival.
Winter Temperatures
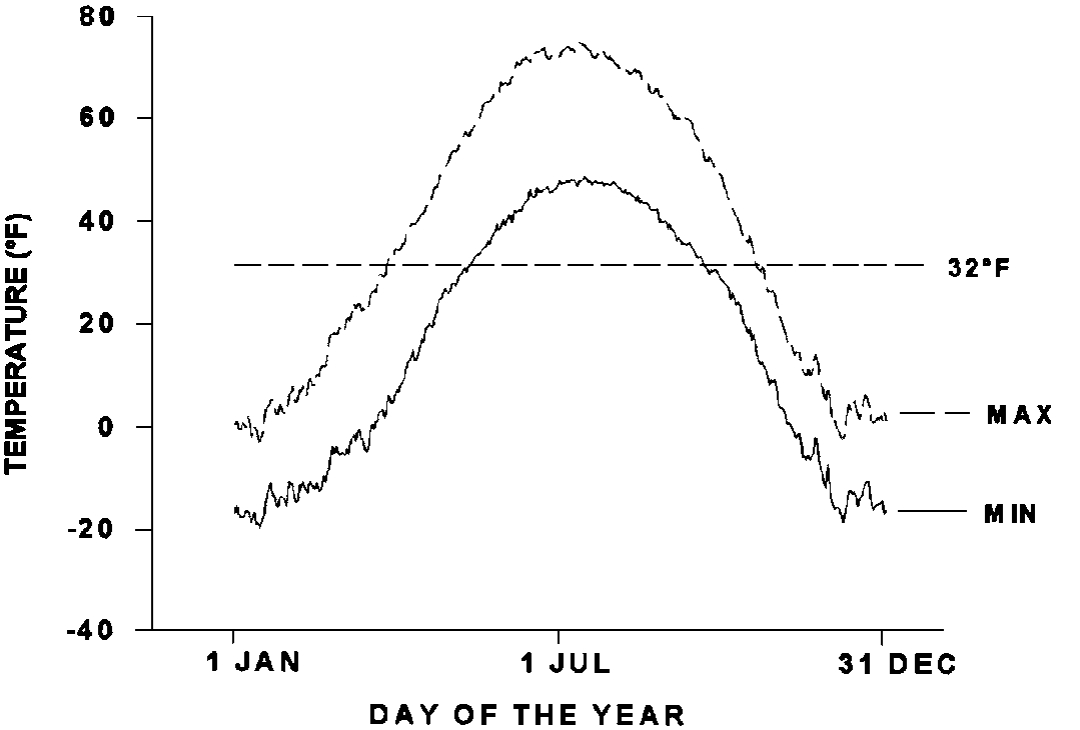
Temperature is one of the first factors to come to mind when discussing winter in Alaska. The severity of winter temperatures and the length of winter depend on the specific location within the state. In Fairbanks, the daily minimum temperature is above freezing for about 130 days and average low temperatures reach minus 20°F (Figure 1). Plant species vary in their tolerance of low temperatures. Temperatures a few degrees above freezing down to 22°F may injure or kill the succulent tissues of herbaceous plants. Some adapted deciduous forest species have the capacity to supercool and tolerate temperatures of minus 40°F or lower. However, even adapted plants can be injured by temperatures below 10°F (Figure 2). The experiment was designed to evaluate the direct effects of low temperatures, so perennial plant organs were removed from the soil and exposed to the indicated temperatures. The strains depicted in the graph vary considerably in their adaptation. Achenbach and Clair originated at 34° to 42°N and they are not as winterhardy as Pumpelly brome (native to Alaska), Polar (selected in Alaska) and Engmo timothy (from Norway at 70°N). The nonadapted cultivars suffer more injury at all temperatures than adapted strains, but exposure to minus 11°F resulted in significant injury, even for cold tolerant, adapted plants. The data illustrate that the average low temperatures in Fairbanks are potentially lethal to adapted and native grass strains. Many factors are involved in winter survival in the field and external factors such as soil insulation and snow cover can affect survival. The balance of this paper will highlight some of the factors that are most important for evaluation, variety selection, and management of perennial plants.
Stresses of Winter

Plants experience a variety of stresses during winter, including direct exposure to low temperatures, ice sheets, desiccation, fluctuating temperatures, energy drain, and disease (Klebesadel 1974, Pollock et al. 1993, Smith 1964). If the stresses exceed the tolerance of the plant, injury or death will result.
Direct Exposure
Cold tolerant plants have the genetic ability to make chemical changes in their cells and thus prevent ice from forming in the cells. Cold hardiness does have limits, however (see Figure 2). Snow cover provides some insulation from ambient air temperatures (Table 1, Figure 13), so if snow cover is lacking or removed by winter winds, plants are at risk (Klebesadel, 1974).
Ice Encasement
In southcentral Alaska, periodic winter thaws will melt snow and water collects in depressions of the fields (Klebesadel 1974). The return to cold weather usually results in formation of ice sheets. Plants may be dormant, but the cells are still alive. As a result, the plants use energy to stay alive and they produce gaseous wastes from life processes. A sheet of ice causes these wastes to accumulate and plants can be killed from the toxic effects of the gases. Small breaks in the ice (from grass stems for example) can greatly reduce damage from ice sheets by allowing some of the gases to escape. Because ice is a better conductor of heat than is snow, plant parts in contact with ice may suffer low temperature damage similar to completely exposed plants as air temperatures drop.
Desiccation
Winter air is dry because cold air cannot hold as much moisture as warm air. Dry winter winds may remove snow and, once plants are exposed, the wind will remove moisture from plant organs near the soil surface. Living cells require some moisture. The amount of moisture needed depends on whether the plant is actively growing or dormant and whether the plant has made chemical changes to reduce freezing damage. During cold hardening, plants remove all unnecessary water from the cells to reduce the possibility of ice forming inside cells. Since virtually all of the unnecessary water has already been removed, dry winds may remove water that is vital for life processes to continue.
Lack of Energy Reserves
Alaskan winters are long and living plants must have sufficient energy stored to sustain life until photosynthesis begins again. The length of winter can be a problem if plants have not been able to store enough reserves. Maintenance of cold hardiness also requires energy. Reserves decline steadily during the winter and mid-winter or early spring thaws followed by a hard freeze can be devastating. Plants are unable to maintain high levels of cold hardiness and may be injured. Poor spring growth may be a symptom of this type of winter injury because plants with low energy reserves cannot replace damaged tissues and sustain vigorous spring growth.
Disease
Diseases, such as snow mold, can also weaken or kill plants. Resistant varieties have been developed for some species and fungicides have been used to control the disease (Goplen and Gossen 1994, Kallio 1966). This problem is not a direct effect of winter in Alaska and it won’t be discussed in detail, but it can further weaken plants (during years when conditions favor disease development) at a time when reserves and cold hardiness are already at low levels.
Importance of Latitude of Origin
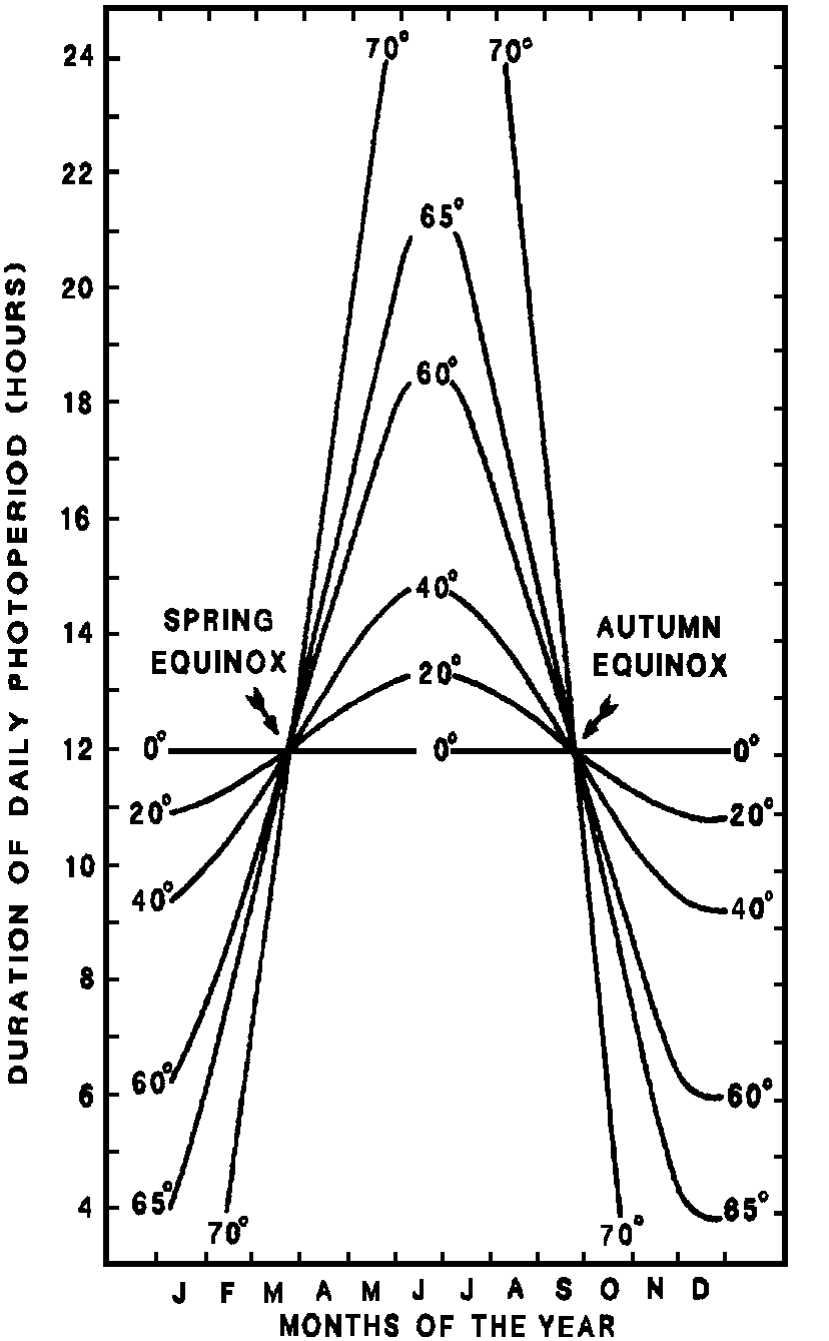
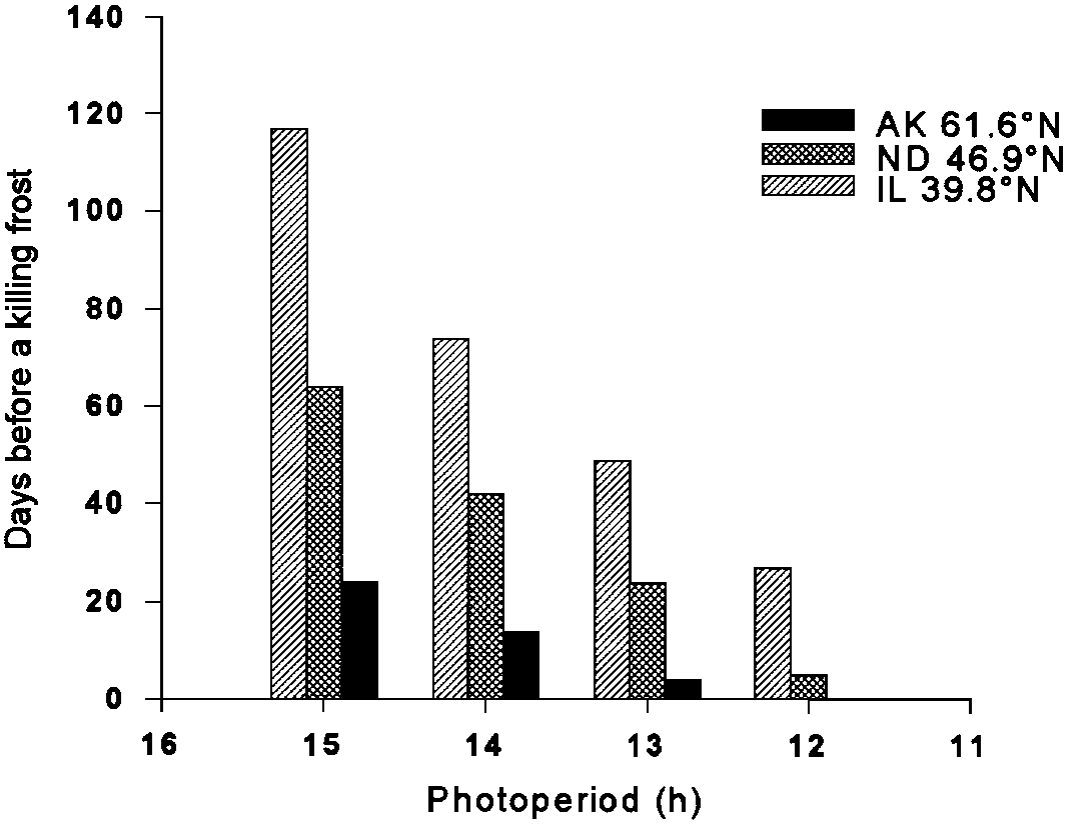
The short growing season in Alaska coupled with the rapid onset of winter make timely preparations for winter vital to plant survival. This raises the question as to how plants are caused to become dormant. The information must be reliable and it must allow for unusual weather patterns. Photoperiod and air temperatures are two of the main signals that plants use as a clock/ calendar. Photoperiod on a given date varies greatly with latitude (Figure 3), but it is consistent from year to year at a given latitude. The signal provided by daylength differs markedly with latitude (Figure 4). A 15 hour photoperiod indicates that a killing frost can be expected in 4 months in Illinois, while the same signal points to a killing frost in 3 weeks in southcentral Alaska. Perennial plants are adapted to interpret the signals at a specific latitude, so latitude of origin is important.
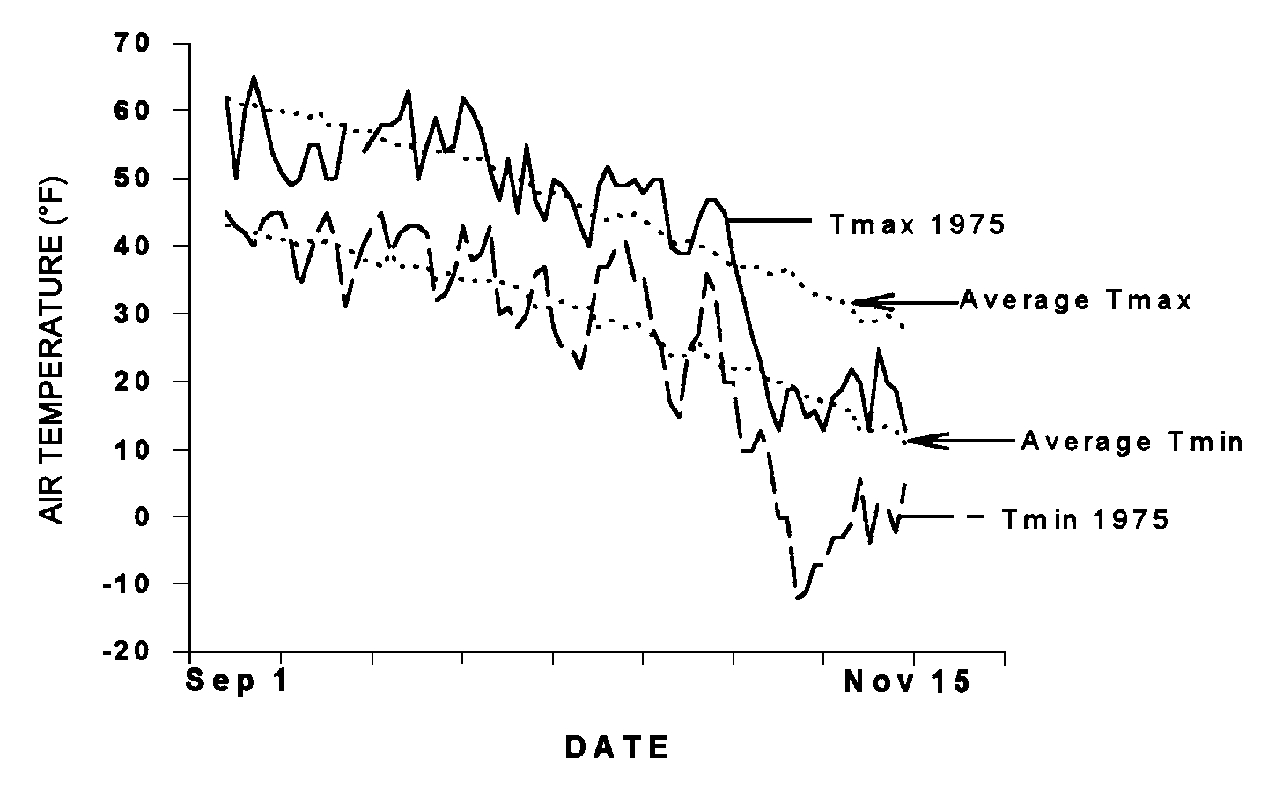
Weather varies considerably from year to year. The daylength provides part of the signal, but the temperature patterns modify the message. Occasionally this causes a problem. An unseasonably warm period was recorded between 10 October and 25 October in 1975 (Figure 5). This period is important for plants because the lowering temperatures promote cold hardiness development. The warm temperatures favored growth over physiological changes that contribute to winter survival. As a result, when the unseasonably cold temperatures hit after 25 October, the plants were not adequately hardened. In spring of 1976, extensive winterkill of both herbaceous and woody plants was observed and it was attributed to the unusual temperature pattern during the previous fall (Klebesadel 1977).
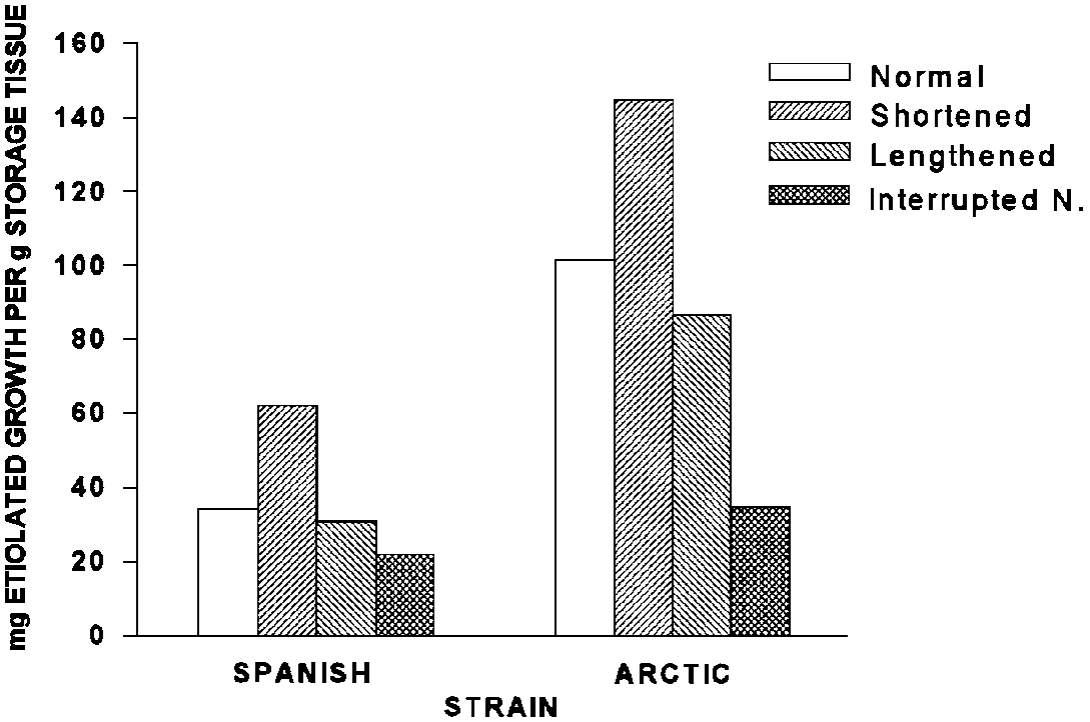
The effects of daylength and temperature in relation to winter survival have been defined for some species. In experiments with sweetclover, Klebesadel (1993c) found that the length of the night was more important than the daylength (Figure 6). The two strains depicted in Figure 6 have different capacities to prepare for winter, but they responded in a similar manner to the treatments. After the treatment period, plants were grown in the dark until no further growth was observed. The weight of the topgrowth produced in the dark (etiolated growth) is a measure of the reserves that the plant had stored. A short day (10.5 to 9 hour photoperiod, long night) simulated autumn conditions and promoted storage of reserves compared to a normally shortening photoperiod (15 to 10.5 hour photoperiod). A long day (15 hour photoperiod) simulated late summer conditions, thus indicating to the plant that winter preparations were not immediately necessary. The interrupted night (normal night 9 to 13.5 hour night) was imposed by supplying 1.5 hour of artificial lighting in the middle of the night. The effect caused by interrupting the night was more dramatic than lengthening the day because the plants use the dark period as a clock. The interruption of the night ‘reset the clock’ and caused the plants to respond as if the night were much shorter and reserve storage was low. Note that the Spanish strain (adapted 35° to 50°N) produced less etiolated growth than Arctic (adapted 50° to 56°N) in response to all treatments.
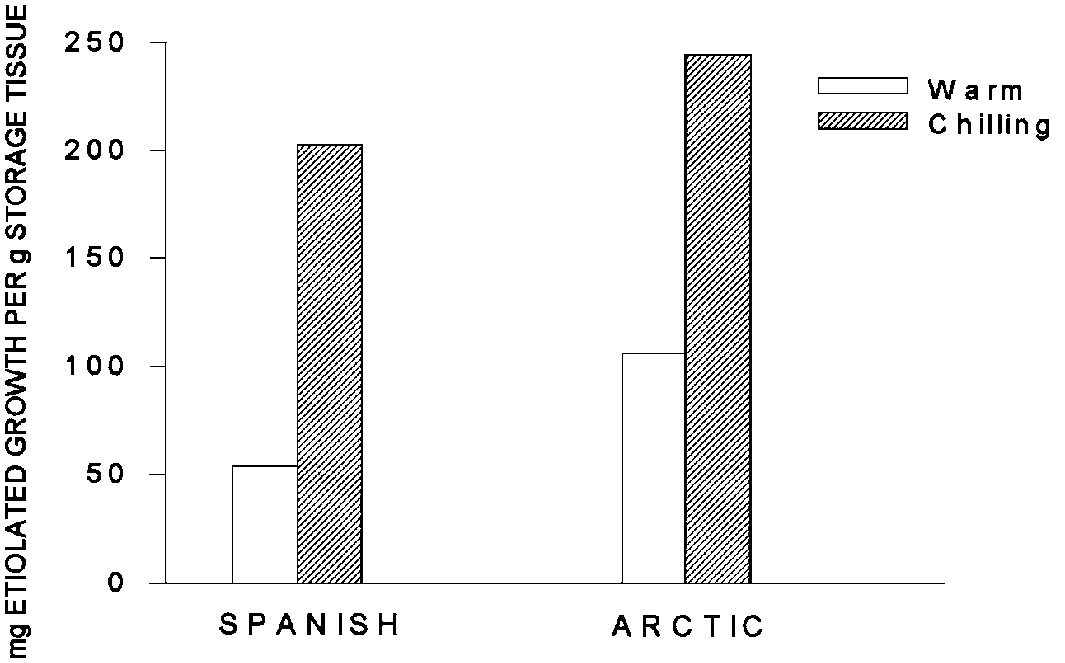
The data in Figure 7 illustrate the importance of temperature in modifying plant response to photoperiod (Klebesadel 1993c). During this experiment photoperiods were shortened while temperatures were either maintained in the 60s or gradually chilled from 40° to 32°F. The warm temperatures discourage storage of reserves (measured by etiolated growth) that are promoted by shortening daylength. The decreasing temperatures increase reserve storage. Note that, as in the previous example, Spanish responds less to the treatments than the more northern-adapted Arctic.
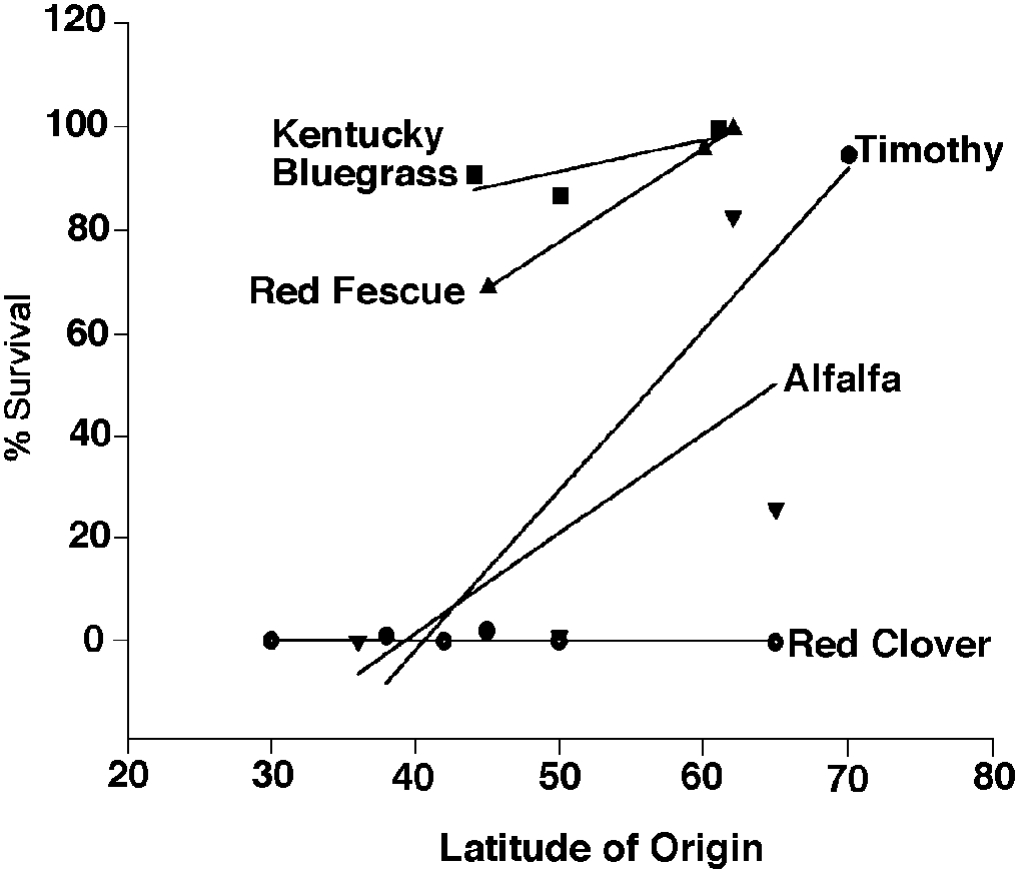
Latitude of origin is extremely important if long term survival is expected because plants must be attuned to the environmental signals (daylength and temperature) that define our seasons. The relative importance of latitude of origin may differ among plant species (Figure 8). The lines describe the average response of a species and the steeper the line, the greater the effect of latitude of origin on survival. Note that for species with the highest survival, latitude of origin s very important. Red clover is one of the least winterhardy species studied and its survival was unaffected by latitude of origin because conditions for this trial were severe and all of the varieties died. At least one variety for each of the other species was capable of surviving.
Mechanisms to Avoid Injury
Energy Reserves
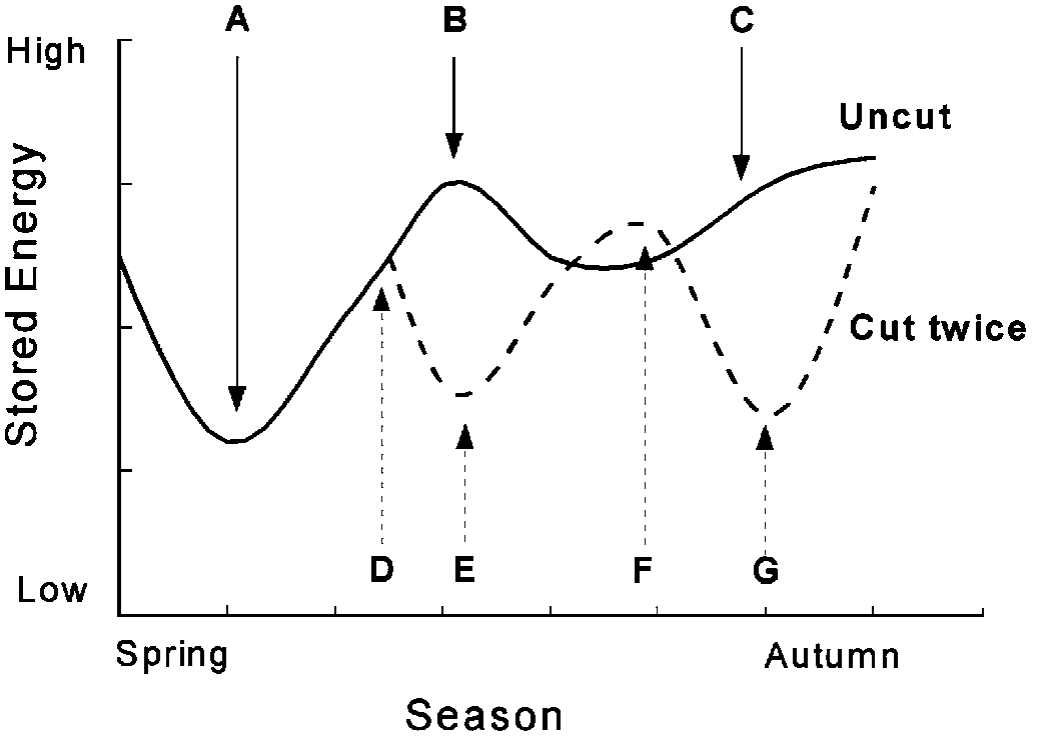
Energy reserves are essential to perennial plants and a good understanding of reserve storage and utilization is useful when making plant management decisions. Two examples of reserve patterns are illustrated in Figure 9. The solid line represents the reserves of an uncut red clover plant. In the spring, reserves are used to support the initial growth (up to A). After a period of reserve-supported growth, the plant has sufficient vegetation to supply energy (from photosynthesis) for current growth and to replace the reserves (A to B). An uncut plant will begin to form seeds and reserves will decline slightly (B to C) and then rise to maximum levels again before winter (from C onward). (NOTE: All plant species do not follow the same patterns of reserve storage and utilization. Differences in plant growth and reproductive development often cause differences in reserve patterns.)
The same plant has a very different reserve pattern when a two cut harvest system is imposed (dotted line). Up to point D the trend is the same as the uncut plant. At D the forage is cut and reserves drop (D to E) as the plant uses stored energy for regrowth following cutting. Reserves are restored (E to F) when the plant has sufficient vegetation to provide for the energy needs of growth and restoration of reserves. At F, the plant is cut again and the reserves fall in the early stages of regrowth (F to G). If adequate time remains in the season, reserves will be restored to high levels before winter. Harvesting too frequently will not allow plants the time to recover before winter. This will increase the likelihood of winter injury. One of the most damaging situations occurs when management causes reserves to decline (F to G), but a killing frost occurs before restoration can take place. Adapted varieties may respond to the shortening days by minimizing regrowth and increasing reserve storage, whereas nonadapted varieties may deplete reserves in regrowth because they do not respond to shortening photoperiods as quickly as adapted plants.
Dormancy
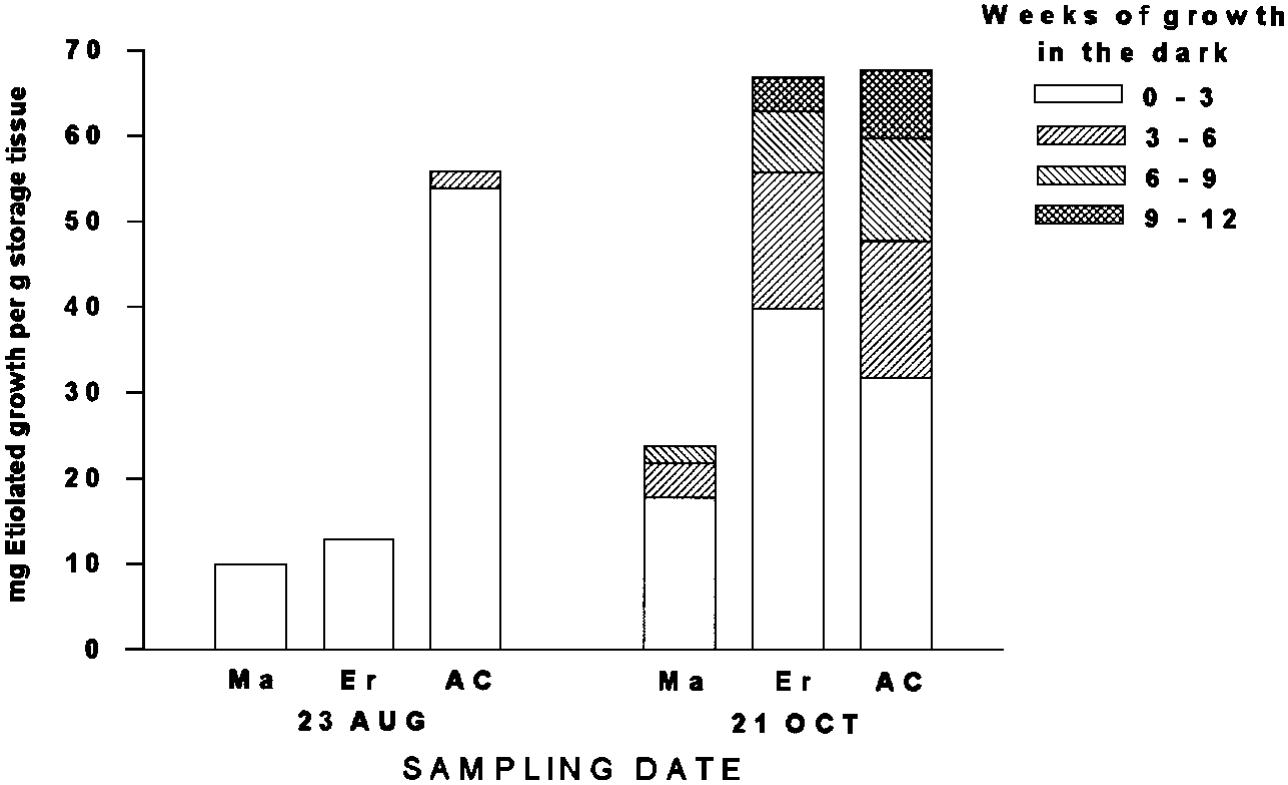
Varieties differ in their capacities to store reserves for the winter period (Figures 6 and 7). In addition, some varieties become more dormant than others. Dormant varieties are less likely to lose cold hardiness and begin growth in response to a mid-winter thaw or a premature breakup. One way to measure dormancy is to determine the amount of etiolated growth (reserves) produced over time. A highly dormant cultivar will continue to produce etiolated growth for longer than a nondormant cultivar. The data in Figure 10 illustrate responses of three strains: Madrid (MA - 40.5°N), Erector (ER- 49.8°N), and Arctic Circle (AC - 66.6°N). All three strains increased reserve storage (measured by etiolated growth) between 23 August and 21 October, but the most northern adapted strain (AC) had a high level of reserves at the earlier date. In August, dormancy was low and virtually all of the reserves were expressed in the first three weeks of growth in the dark. By October, the plants had become more dormant and the length of time required to totally deplete reserves increased. Note that as latitude of origin increased, a higher proportion of the reserves were expressed in later regrowth periods. This indicates that the most northern adapted strain (Arctic Circle) is the most dormant of the three.
Hardening
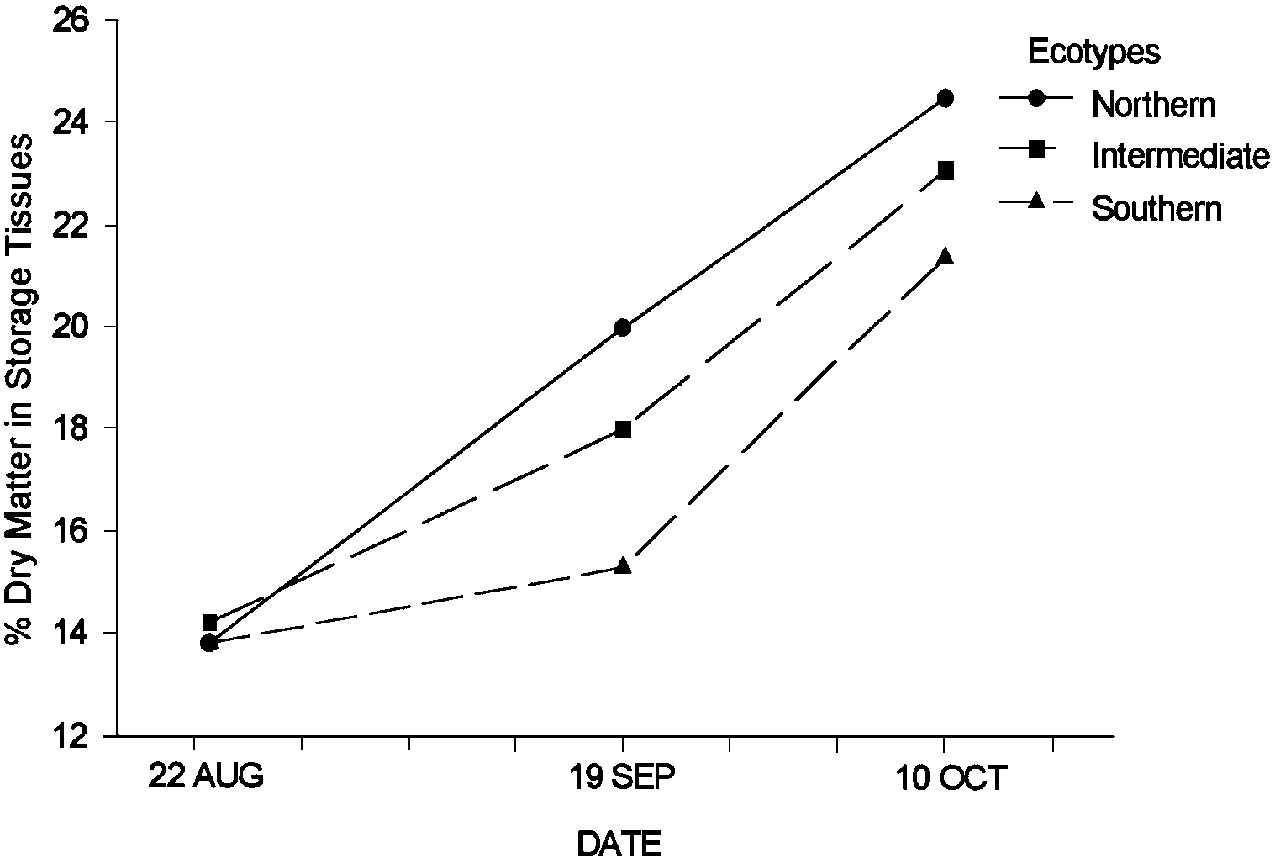
Plants undergo many physiological changes when they harden. One indication of hardening is an increase in the percent dry matter of perennial organs. Reserves are stored in the cells and free water is removed to reduce the potential for damage from sub-freezing temperatures. Both of these changes increase the dry matter concentration in perennial organs. Plants that are adapted to northern latitudes tend to begin to make these physiological changes for winter earlier than nonadapted plants (Figure 11). In addition, northern ecotypes often have greater genetic capacity than southern ecotypes to store reserves and to maintain a hardened condition and thus they are more capable of surviving the long winters at this latitude. The data in the graph include a wide range of strains and species. It is clear that differences in timing and extent of winter preparations are typical of ecotypes that are adapted to survive at different latitudes.
Morphology
In addition to latitude of origin, plant morphology plays a role in the ability of a plant to survive harsh winter conditions (Klebesadel 1994a). Plants that have their perennial organs well underground are more insulated from direct exposure to low temperatures. The soil provides some insulation and a snow cover further protects the plants. Rhizomatous species also have the capacity to ‘fill in’ and compensate for injury when necessary. Plants that have perennial organs at or above the soil surface are subject to winter injury when exposed to low temperatures before snow cover develops, during a midwinter thaw, or following an early spring snow melt. Desiccation from winter winds can also be a problem if the winds have removed the protective snow cover.
Complicating Factors
Management Practices
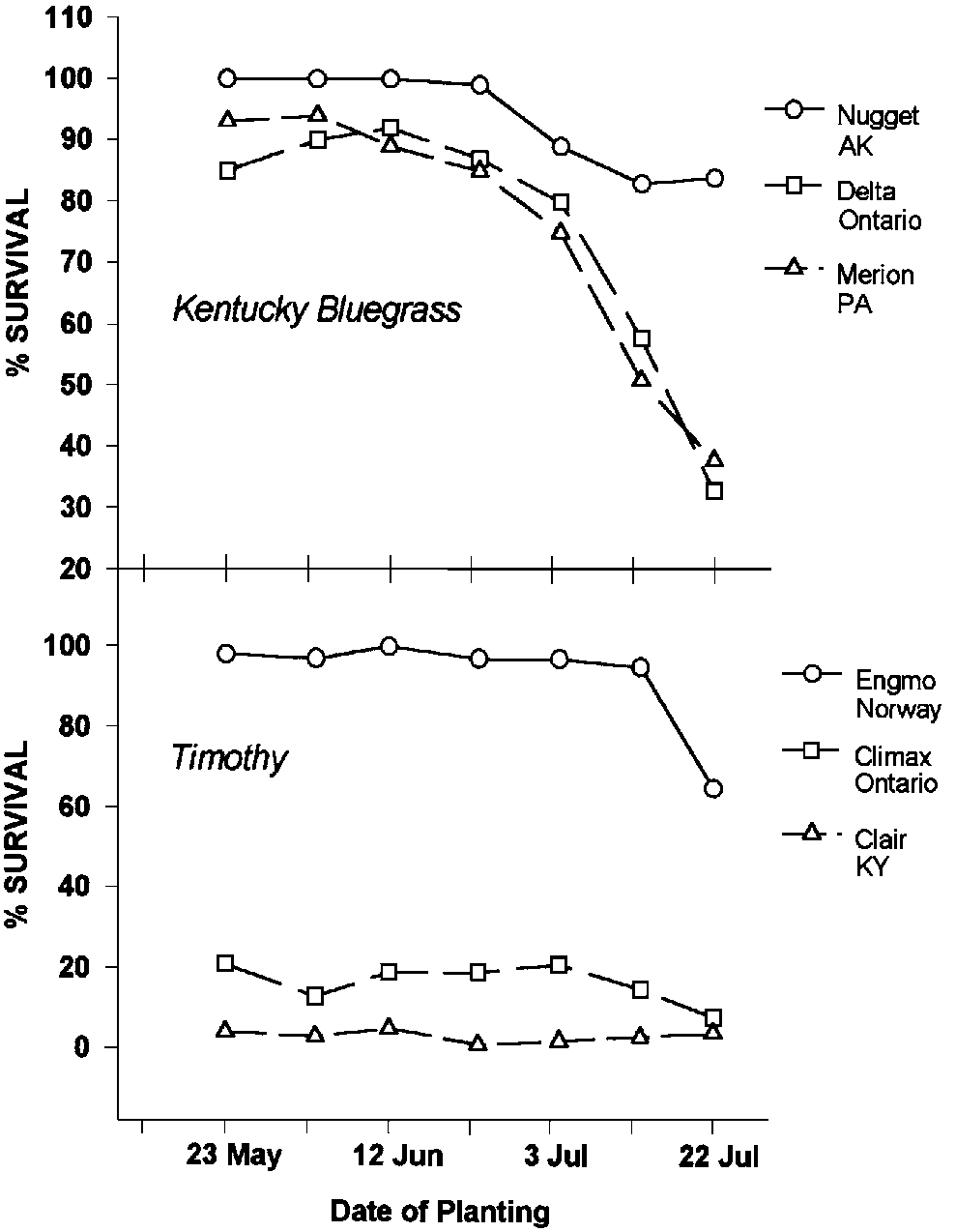
Many management practices can affect winter survival of perennials. Fertilization, planting dates, disease and insect pests, cutting dates, cutting frequency, and other factors can enhance or reduce survival of perennial plants. The data in Figure 12 illustrate the effect of planting dates on survival of six grass varieties. The most northern adapted Nugget and Engmo have a higher percent survival and they tolerate a wider range of planting dates compared to the other varieties. It should be noted that even the hardiest plant can be predisposed to winter injury by management. Klebesadel (1994b) studied the responses of Polar brome to cutting management. Polar is normally very winterhardy, but the combination of three cuttings followed by a severe winter resulted in a 59% winterkill of Polar. Winterkill was 16% in Polar stands that had only been harvested twice.
Snow
Snow is a very effective insulator. The rate of heat transmission through a substance (thermal conductivity) is used to define the insulating properties of that substance. The data in Table 1 indicate that snow is quite effective as an insulator when compared to water, ice, or iron. The practical value of snow can best be seen by looking at soil temperatures at sites that differ in snow cover. The Matanuska Farm near Palmer is in a location that is strongly affected by winter winds. Snow cover is periodically removed during the winter leaving little protection for the perennial plants (Klebesadel 1974). Point MacKenzie is located about 28 miles southwest of the Matanuska Farm. While the air temperatures are comparable at these two locations, the snow cover is very different because the winds that remove snow in the Palmer area do not affect Point MacKenzie. The soil and air temperatures for the two sites are presented in Figure 13. The insulating qualities of snow have prevented soil temperatures at Point MacKenzie from going below 29°F for the past four winters, while soil temperatures regularly fall to between 0° F and 10°F at the Matanuska Farm. The protection of snow is so effective that poorly adapted varieties of red and white clover (from Florida and California) have survived recent winters. Good snow cover can mask deficiencies in winterhardiness of poorly adapted varieties. Any combination of events that results in a lack of snow cover together with low temperatures will expose the limitations.
Year-to-Year Weather Variation
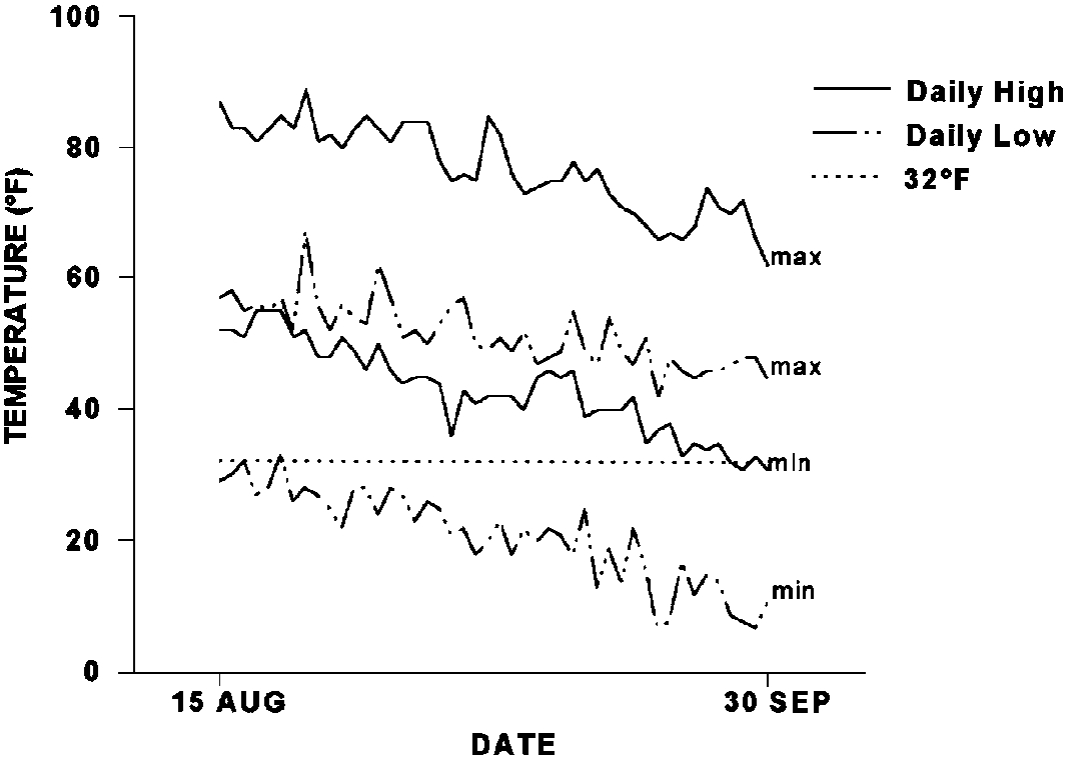
The weather never seems to be ‘average’. Year- to-year variations can be large. Differences in temperature extremes, precipitation, and timing combine to make winters that are described as early, late, hard, mild, open, etc. Figure 14 illustrates the variation in high and low temperatures for the critical period during which perennial plants are storing reserves and making some of the other changes associated with winter survival. At the end of September, daily high temperatures will be somewhere between 31° and 62°F, while the daily low will be between 11° and 45°F. This wide range indicates why plants must adjust the daylength signal. With a short growing season, plants must maximize growth in order to compete effectively. Yet they must also be prepared to become dormant early when the growing season ends early. Adapted plants effectively make small adjustments to make the best use of the growing season and they compensate for minor changes in reserve levels, cold hardiness, etc. that result from minor year-to-year variations. The more drastic departures from the normal trend can have catastrophic results as noted earlier (Figure 5) or misleading results. Mild winters and early, deep snow cover can hide deficiencies in winterhardiness. Evaluations should be made over several winters to reliably rank the hardiness of varieties.
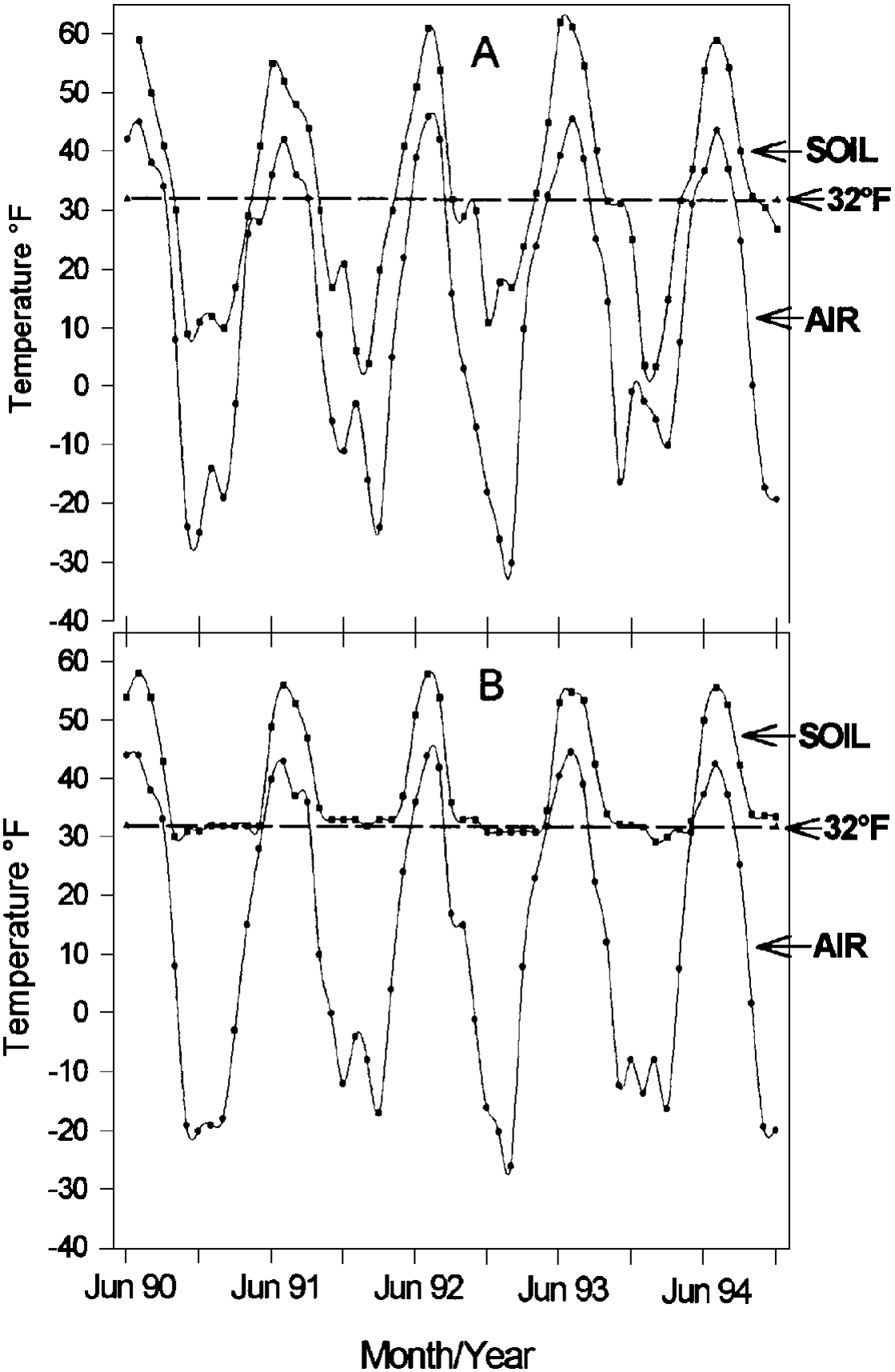
Table 1. Quantity of heat (calories) transmitted per second through several substances
| Substance | Thermal Conductivity (10 –4 ) |
|---|---|
| Air | 0.57 |
| Dry soil | 3.30 |
| Compact snow | 5.10 |
| Water | 13.48 |
| Ice | 50.00 |
| Iron | 16.10 |
Source: Smith et al., 1986
Test Conditions
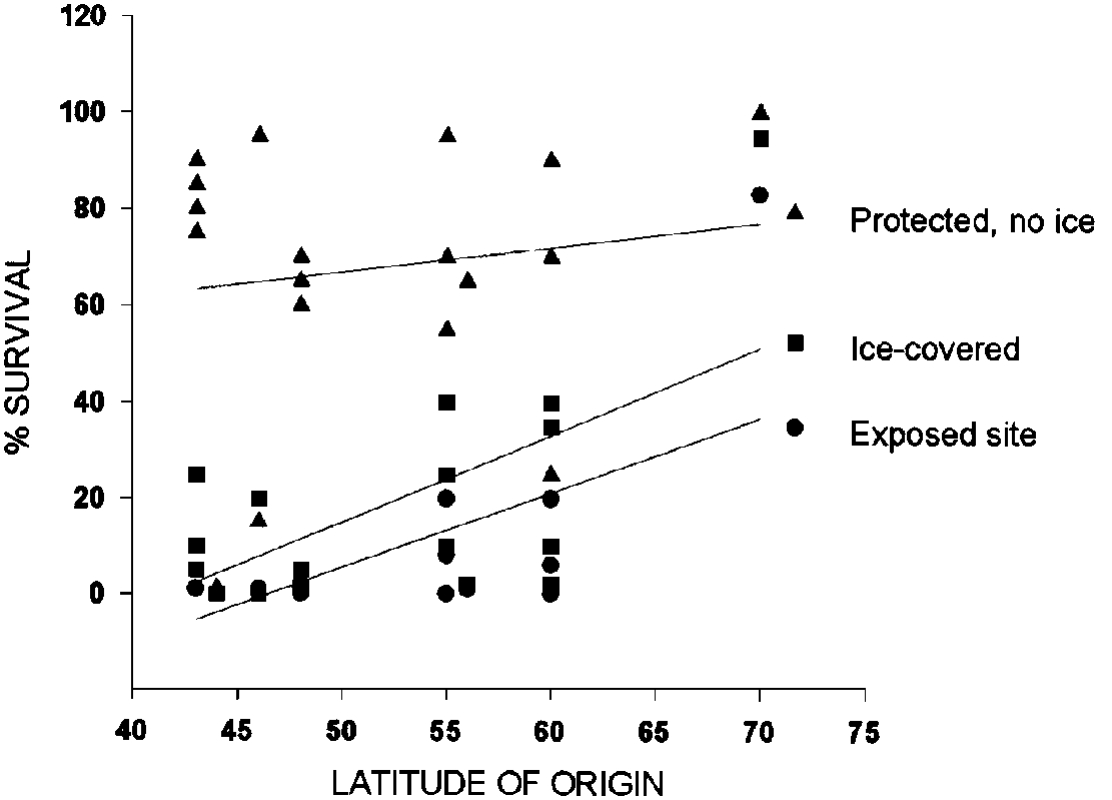
We have been discussing the importance of latitude of origin for proper development of winterhardiness. The plants must be able to understand the environmental signals that mark the changing seasons. However, someone is always importing a plant that shouldn’t survive. Why does it work for some and not for others? The data in Figure 15 illustrate one way that this can happen. Under certain conditions, (protected site, no ice) the latitude of origin seems to have little to do with survival. Some of the plants that originated at 40° to 45°N survive as well as those originating at 60°N. When conditions become severe (ice or exposed with snow removal), latitude of origin is very important to survival. Poorly adapted plants will survive Alaska winters if they are grown in protected sites. Variable success with a variety or species in the same part of the state suggests that site is an important factor.
Plant Hardiness Zones
Plant hardiness zone maps are based primarily on temperatures. As much of Dr. Klebesadel’s work demonstrates, temperature tolerance alone is not a good measure of adaptation to Alaska growing conditions. If plants do not respond to the environmental signals that mark the changing seasons, they may not be able to demonstrate the ability to tolerate low temperatures.
Summary
Perennial plants in Alaska are exposed to a variety of stresses during the winter: low temperatures, fluctuating temperatures, desiccation, long dormant period, ice sheets, and disease. If these plants are to survive, they must store an adequate amount of reserves, develop internal cold hardiness, and become dormant. Adapted plants use signals from daylength and ambient temperature to initiate winter preparations. The ability to recognize the daylength signal is related to the latitude of origin for a variety. Northern adapted varieties generally begin to make the necessary changes earlier than varieties from lower latitudes. In addition to the above physiological adjustments for winter, plant morphology and snow cover can be important factors in plant survival.
When perennial plant parts are protected by the soil and snow, survival is enhanced.
Using This Information
There are at least two ways to use the information presented above:
Minimize Risk: Use and recommend only adapted plants. Avoid importing plants from lower latitudes. This approach will limit winter-kill losses. It should be noted, however, that some winters are so severe that even adapted plants (selected and tested in Alaska) will winterkill.
Accept Risk: Many plants are capable of surviving most winters if planted in a protected site and carefully managed. Some individuals are willing to accept periodic losses of perennial plants in return for the aesthetic or economic benefits of particular plant species. These people should be encouraged to experiment with different species and cultivars, but before they are encouraged, they should understand the nature of winter survival and the value of protected sites.
Acknowledgments
While it is evident from the references cited, the author would like to further emphasize Dr. L. J. Klebesadel’s important contribution to our understanding of coldhardiness and perennial plant survival at extreme northern latitudes. I also thank Dr. Klebesadel for his careful review of this manuscript.
References
Goplen, B. P. and B. D. Gossen. 1994. Cultivar description: AC Nordica alfalfa. Canadian Journal of Plant Science 74:145–147.
Kallio, A. 1966. Chemical control of snow mold (Sclerotinia borealis) of four varieties of bluegrass (Poa pratensis) in Alaska. Plant Disease Reporter 50:69–72.
Klebesadel, L. J. 1974. Winter stresses affecting overwintering crops in the Matanuska Valley. Agroborealis 6(1):17–20.
Klebesadel, L. J. 1977. Unusual autumn temperature pattern implicated in 1975–76 winterkill of plants. Agroborealis 9(1):21–23.
Klebesadel, L. J. 1985. The critical importance of north-latitude adaptation for dependable winter survival of perennial plants in Alaska. Agroborealis 17(1):21–30.
Klebesadel, L. J. 1992a. Effects of Planting Date and Latitude-of-Adaptation on Seeding-Year Development, Winter Survival and Subsequent Seed and Forage Production Potential of Grasses and Legumes in Alaska. University of Alaska, Agriculture and Forestry Experiment Station. Bulletin No. 86.
Klebesadel, L. J. 1992b. Extreme Northern Acclimatization in Biennial Yellow Sweetclover (Melilotus officinalis) at the Arctic Circle. University of Alaska, Agriculture and Forestry Experiment Station. Bulletin No. 89.
Klebesadel, L. J. 1992c. Morphological, Physiological and Winterhardiness Comparisons Among Latitudinal Ecotypes of Biennial Sweetclover (Melilotus species) in Subarctic Alaska. University of Alaska, Agriculture and Forestry Experiment Station. Bulletin No. 91.
Klebesadel, L. J. 1993a. Bromegrass in Alaska II. Autumn Food-Reserve Storage, Freeze Tolerance and Dry-Matter Concentration in Overwintering Tissues as Related to Winter Survival of Latitudinal Ecotypes. University of Alaska, Agriculture and Forestry Experiment Station. Bulletin No. 93.
Klebesadel, L. J. 1993b. Winter Survival of Grasses and Legumes in Subarctic Alaska as Related to Latitudinal Adaptation, Pre-winter Storage of Food Reserves and Dry-Matter Concentration in Overwintering Tissues. University of Alaska, Agriculture and Forestry Experiment Station. Bulletin No. 94.
Klebesadel, L. J. 1993c. Effects of Daily Photoperiod/Nyctoperiod and Temperature on Autumn Development of Crown Buds and Dormancy, Freeze Tolerance and Storage of Food Reserves in Latitudinal Ecotypes of Biennial White Sweetclover. University of Alaska, Agriculture and Forestry Experiment Station. Bulletin No. 95.
Klebesadel, L. J. 1994a. Winterhardiness, Forage Production and Persistence of Introduced and Native Grasses and Legumes in Southcentral Alaska. University of Alaska, Agriculture and Forestry Experiment Station. Bulletin No. 101.
Klebesadel, L. J. 1994b. Bromegrass in Alaska IV. Effects of Various Schedules and Frequencies of Harvest on Forage Yields and Quality and on Subsequent Winter Survival of Several Strains. University of Alaska, Agriculture and Forestry Experiment Station. Bulletin No. 102.
Klebesadel, L. J. and S. M. Dofing. 1991. Reed Canarygrass in Alaska: Influence of Latitude-of-Adaptation on Winter Survival and Forage Productivity and Observations on Seed Production. University of Alaska, Agriculture and Forestry Experiment Station. Bulletin No. 84.
Klebesadel, L. J. and D. Helm. 1986. Food reserve storage, low-temperature injury, winter survival and forage yields of timothy in subarctic Alaska as related to latitude-of-origin. Crop Science 26:325334.
Panciera, M. T. and S. D. Sparrow. 1994. Guidelines on Tolerances, Limitations and Risks Associated with the Use of Forage Legumes in Alaska. Alaska Cooperative Extension Crop Production and Soil Management Series No. FGV-00247A.
Pollock, C. J., C. F. Eagles, C. J. Howarth, P. H. D. Schunmann and J. L. Stoddart. 1993. Temperature stress. In Plant Adaptation to Environmental Stress, edited by L. Fowden, T. Mansfield and J. Stoddart. London: Chapman and Hall Ltd.
Smith, Dale. 1964 Winter injury and the survival of forage plants. Herbage Abstracts 34:203–209.
Smith, Dale, R.J. Bula and R.P. Walgenbach. 1986. Forage Management in the North. 79–90. Dubuque, Iowa: Kendall/Hunt Publishing Co.
Julie Riley, Extension Faculty, Agriculture and Horticulture. Originally prepared by Michael T. Panciera, former Forage Agronomist, Agricultural and Forestry Experiment Station, University of Alaska Fairbanks
Revised March 2023